2.カルニチンの役割
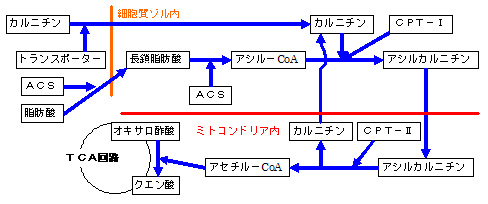
a.カルニチンは、脂肪酸を、ミトコンドリア内に輸送するのに必要
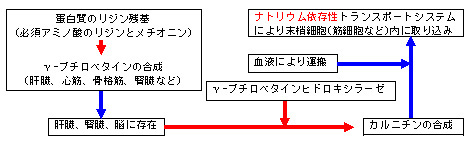
・ カルニチンは、カルニチントランスポーターにより、細胞内に取り込まれます。
・ 細胞質の長鎖脂肪酸は 、細胞質ゾルで、ACS(Acyl-CoA synthase)により、CoA(Coenzyme A)と結合して、アシル-CoA(Acyl-CoA)に活性化 されます。
・ アシル-CoAは 、CPT-I(carnitine O カルニチンと結合 して、アシルカルニチン(Asylcarnitine)となって、ミトコンドリアの内膜を通過し、ミトコンドリア内のマトリックスに移行します。従って、カルニチンは、ミトコンドリア外の長鎖 脂肪酸を、ミトコンドリア内に輸送する のに必須な物質(ビタミン)であり、カルニチンがないと、長鎖 脂肪酸は、ミトコンドリア内のマトリックスで、β-酸化 されにくくなります。ミトコンドリア内のマトリックスに入ったアシルカルニチンは、CPT-II(carnitine O
・ アシル-CoAに活性化された長鎖脂肪酸は、β-酸化によりアセチル-CoAに変換 されオキサロ酢酸と結合してTCA回路(クエン酸回路)に取り込まれます。
注 肝臓では、遊離脂肪酸から生成されるアシル-CoAが、カルニチンシャトルにより、ミトコンドリア内に取り込まれ、アセチル-CoAとなり、TCA回路で代謝されます。インスリンは、このアシル-CoAのミトコンドリア内への取り込みを抑制します。インスリン不足・インスリン拮抗ホルモン過剰 の際には、アシル-CoAのミトコンドリアへの取り込みが促進され、大量のアシル-CoA(遊離脂肪酸から生成される)が、ミトコンドリア内に流入 し、TCA回路で代謝されないと、アセチル-CoAから、ケトン体が生成 され、血中ケトン体が増加し、ケトーシスを来たします。
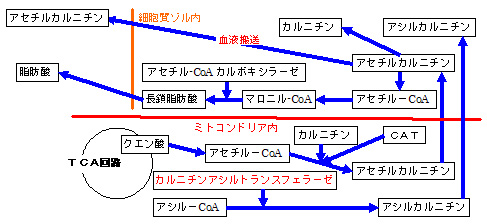
b.カルニチンは、脂肪酸を、ミトコンドリア外に輸送するのに必要
・ ミトコンドリア内のアセチル-CoA は、カルニチンアセチルトランスフェラーゼ(カルニチンと結合 して、アセチルカルニチンとして、ミトコンドリア外の細胞質ゾルに輸送 されます。 ア セチルカルニチンは、細胞質ゾルから、血液中に排泄されます。また、細胞質ゾル内のアセチルカルニチン は、カルニチンとアセチル-CoAに分解 され、アセチル-CoAは 、アセチル-CoAカルボキシラーゼ(acetyl-CoA carboxylase:ACC)により、マロニル-CoA経路で、脂肪酸に合成 されます。
・ 脂肪酸アシル-CoA(LCACoA)がミトコンドリア内に蓄積 すると、ミトコンドリア内膜に存在するカルニチン の作用により、活性脂肪酸(acyl-CoA)のアシル基(acyl groups)を、カルニチン(L-carnitine)に転移させ、ACR(アシルカルニチン)を生成することで、、ミトコンドリア外に輸送させます。
・ カルニチンは、ミ トコンドリア内のアセチル-CoAや、アシル-CoAを、ミトコンドリア外に輸送 する(ミトコンドリア内のCoA/アシル-CoA比を調節する)のに必要な物質でもあります。
注
・
カルニチンアセチルトランスフェラーゼ(carnitine O カルニチンアシルトランスフェラーゼ であり、acetyl-CoA(C2 C3 C4 acetyl-CoA + carnitine = CoA + O- acetylcarnitine
・ カルニチンパルミトイルトランスフェラーゼ(carnitine O アシルトランスフェラーゼ であり、炭素数が8〜18(C8 〜C18 C16 )の活性脂肪酸(palmitoyl-CoA)に最も作用します。palmitoyl-CoA + L-carnitine = CoA + L-palmitoylcarnitine
・ カルニチンオクタノリルトランスフェラーゼ (carnitine O -octanoyltransferase:EC 2.3.1.137.)は、中鎖〜長鎖(medium-chain/long-chain)の活性脂肪酸(acyl-CoAs)に作用 し、特に、炭素数が6〜8(C6 〜C8 )のアシル基(acyl groups)のオクタン酸(C8 )の活性脂肪酸(octanoyl-CoA)などを始めとして、作用します。octanoyl-CoA + L-carnitine = CoA + L-octanoylcarnitine
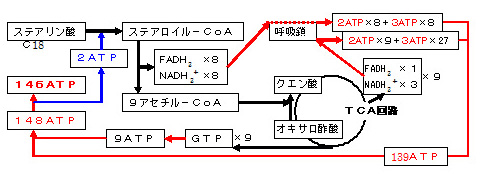
c.カルニチンをエネルギー産生から見た場合
・ カルニチンによってミトコンドリア内へ輸送された長鎖脂肪酸のβ-酸化によるアセチル-CoAの生成 は、エネルギー産生 につながるだけでなく、ピルビン酸脱水素酵素を抑制 し、解糖(グルコースのピルビン酸への分解)が抑制され、ピルビン酸からの乳酸生成が減少 します。しかし、低酸素(hypoxia)などによるTCA回路(クエン酸回路)の障害 ・脂肪酸の過剰摂取 や脂肪酸からのエネルギー産生を必要としない 場合、余剰となるアセチル-CoAは カルニチンによって、アセチルカルニチンに変換されたり、マロニル-CoA経路で再び脂肪酸に合成 されます。これにより、アセチル-CoAによるピルビン酸脱水素酵素の抑制は軽減 されます。
・ 炭水化物の過剰摂取 は、ピルビン酸脱水素酵素により、ピルビン酸からアセチル-CoAが生成 され、アセチルカルニチンとして、細胞質ゾルに輸送され、アセチル-CoAから、アセチル-CoAカルボキシラーゼにより、マロニル-CoA経路で再び脂肪酸に合成 されます。マロニル-CoAは、CPT-Iを抑制 し、脂肪酸のβ-酸化を抑制 します。( 糖質の代謝参照)
a.カルニチンの代謝
・ カルニチンの体内総プール量は、100mmolであり、その内、98%が筋肉内 に、1.6%が肝臓と腎臓に、0.6%が細胞外液中に存在します。
カルニチンは、大半は尿中(腎臓)から排泄 され、1%以下は、便中から、排泄されます。
・ 腎臓でのクリアランスは、アシルカルニチンは遊離カルニチンより、10〜20倍速い。アセチルカルニチンは、腎尿細管から再吸収されますが、長鎖脂肪酸のアシルカルニチンは、再吸収が悪い。その為、脂肪酸代謝異常症(FAOD) では、代謝が阻害された脂肪酸の炭素鎖が長い程、アシルカルニチンとして、カルニチンが、尿中に排泄 されるので、血中のカルニチンは減少 し易い。
・ 尿中に排泄されるアシルカルニチンを分析することにより、脂肪酸代謝異常症(FAOD)や、有機酸代謝異常症を、診断することが可能 です。
・ ライ症候群 でも、ミトコンドリアの脂肪酸代謝(β-酸化)が障害され、アシルカルニチンが、尿中に増加 します。正常児のアシルカルニチンのアシル基は、アセチル基(アセチール)である(アセチルカルニチン)が、ライ症候群のアシルカルニチンのアシル基は、アシル基とされます。また、ライ症候群に類似した症状を示す、カルニチン欠損症と異なり、ライ症候群では、血中カルニチン値は、基本的には低下しませんが、二次的に低下することがあります。
b.慢性疲労症候群(CFS )
カルニチン(アシルカルニチン)が欠乏すると、慢性疲労症候群(CFS) と言う病気(風邪などを契機に、激しい倦怠感や、脱力感を、半年以上、持続ないし繰り返す)になるとも言われています。慢性疲労症候群の病因は確定されていません が、過度なストレスから生体機能を守る為に、自己防衛機制が作動し、コルチゾールやDHEA-Sの産生を抑制したり、脳血流を低下させて脳代謝を抑制したりしていると考えることもできます。
・ ACR(アシルカルニチン)は、 グルタミン酸、GABA(γアミノ酪酸)などの神経伝達物質の合成 に利用されますが、慢性疲労症候群では、ACR(アシルカルニチン)の脳内(前頭葉の前帯状回と、前頭前野)への取り込みが低下していると言われており、ACRは疲労の程度と相関 していると考えられます。また、アセチルカルニチンの脳内への取り込み が、Broadmann24野(自律神経系の調節や情動などに関連する)と、Broadmann9野(意欲やコミュニケーションに関連する)において、局所脳血液量低下とは無関係に、低下している と言われています。慢性疲労症候群では、アシルカルニチンやアセチルカルニチンの不足と脳内への取り込みの低下が同時に起こっていることから、ストレスなどによる体内の代謝の抑制 により、アシルカルニチンやアセチルカルニチンの産生(合成)や、細胞内への取り込みが低下していると仮定することもできる。
・ 慢性疲労症候群では、血清コルチゾールやDHEA-S(デヒドロエピアンドロステロンサルフェート)など、副腎皮質から産生されるホルモンが減少 し、思考力や集中力が低下 すると言われる。DHEA-Sは、アセチルカルニチントランスフェラーゼ(acetylcarnitine
transferase:carnitine O DHEA-Sが減少すると、アセチルカルニチンが低下 する。DHEA-Sを投与(補充)すると、アセチルカルニチンが上昇する。この説では、アセチルカルニチンの低下は、慢性疲労症候群の原因ではなく、結果を意味します 。ストレスにより、内分泌系が崩れ、DHEA-Sなどのホルモン産生のバランスの失調 が、慢性疲労症候群の第一原因 と考えることができる。。
・ 慢性疲労症候群では、NK細胞活性が低下 し、潜伏感染していたウイルス(HHV-6、EBVなど)が、再活性化する と言われ、それに伴い、抗炎症性サイトカインのTGF-βが増加 すします。TGF-βは、DHEAの硫酸抱合を調節している酵素(スルフォキナーゼ)の活性を抑制する為、DHEA-Sが減少 し、アセチルカルニチンの合成が低下 します。また、IFN-α(インターフェロンアルファ)は、2-5AS(2',5'-オリゴアデニル酸合成酵素)を誘導し、誘導された2-5A(2',5'-オリゴアデニル酸)により、RNaseを活性化させ、ウイルスRNAを分解して、ウイルス増殖を抑制するが、慢性疲労症候群では、2-5ASが上昇 している。しかし、HHV-6、EBVなどのウイルスは、初感染後、体内に、潜伏感染するウイルスであり、特に、HHV-6は、過労状態の人では、高率に唾液中に分泌(検出)される。従って、ウイルスの再活性化や、2-5ASの上昇 は、慢性疲労症候群の原因と捉えるより、ストレスにより、免疫力が低下(疲弊)した結果 と、捉える方が、妥当と思われる。
c.全身性カルニチン欠損症
全身性カルニチン欠損症では、カルニチントランスポーターに障害があり、骨格筋、心筋、腎臓で、血液中からカルニチンを、細胞内に取り込めない 。また、血中カルニチンも減少 している。これにより細胞内カルニチンが欠乏するため、長鎖脂肪酸を、ミトコンドリア内へ転送する能力が低下し、脂肪酸は、ミトコンドリアでβ-酸化をされないため、飢餓などで、グルコースからアセチル-CoAを生成できない状況になると、エネルギー産生(ATP産生)が低下 します。また、糖新生も低下して、ケトン体の上昇が少ない、低ケトン性低血糖 を来たします。中性脂肪(トリグリセリド)が組織に蓄積 し、肝腫大、心筋障害、ミオパチーなどを引き起こします。有害なアシル-CoAが蓄積 し、TCA回路(ピルビン酸脱水素酵素)や、尿素回路(N-アセチルグルタミン酸合成酵素)が阻害 され、エネルギー産生の低下、高乳酸血症、高アンモニア血症などを呈します (ライ症候群に類似した症状を来たします)。
d.抗生剤による、カルニチン欠乏
ピ ボキシル基を持つ抗生剤抗菌活性物質とピバリン酸に分解 されます。ピバリン酸は、カルニチン抱合を受け、尿中にピバロイルカルニチンとして排出 されます。従って、ピボキシル基を持つ抗生剤を長期間投与すると、血清中カルニチンが低下 すします。